РЕВМАТОИДЕН АРТРИТ - ПАТОГЕНЕТИЧНИ АСПЕКТИ
Arthrytis Rheumatodes - Pathogenic Aspects
Dr. Kiril Yablanski
Medical University - Pleven
Разглеждайки ревматоидният артрит (РА) в патогенетичен аспект, бихме могли да го дефинираме като автоимунна, извънсъдова, имунокомплексна болест, локализирана в ставните структури. Автоимунн ;ата възпалителна реакция е резултат от взаимодействие между генетични и външни фактори.
Етиологията на РА все още не е известна, но през последните години се натрупа достатъчно фактически материал, даващ основание да се предполага инфекцията като основен причинен момент.
Ставите са обект на инвазия от различни микроорганизми: гонококи, хламидии, борелии, ретровируси, йерсинии. Микроорганизмите могат да участват в патогенезата на РА не само чрез директно проник_ 6;ане в ставите, но и по други механизми:
Съществена роля в патогенезата на РА се пада на вътрешните (генетични) фактори. Те могат да бъдат сведени до два основни - (а) група генетично детерминирани антигени, разположени по клетъчната повъ ;рхност на повечето ядроносни клетки в организма (HLA система) и (б) гените на главния комплекс на тъканната съвместимост (МНС).
РА е първото заболяване, за което е доказано, че асоциира с детерминанти на локус D (DR4, DR1, Dw14)от HLA. Повечето автори свързват носителството на тези антигени с предиспозиция към РА, както и с по-тежко протича_ 237;е на болестта. Хипотезите, обясняващи тази асоциация могат да бъдат представени по следния начин:
Приблизително половината от болните с РА са носители на HLA DR4 антиген, докато при здравите честотата му е около 18%.
От DQ антигените е доказана връзката между РА и носителството на DQ2, който се смята за генетичен маркер за развитието на това заболяване. DQ1 при здрави лица се счита като протективен генетичен фактор _ 1;а развитието на болестта.
В развитието на ревматоидната възпалителна реакция условно могат да се обособят пет фази.
I. Начална фаза
. В нея инфекциозният (вирусен?) агент навлиза в ставата по хематогенен път. Като най-вероятни причинители на РА за сега се считат вирусът на Ebstein-Barr, парвовирусът RA-1 и вирусъ_ 242; на рубеолата. Следва фиксацията на вируса в ставната синовия. Антиген-представящите клетки го инкорпорират в цитоплазмата си, преработват го до отделни пептидни фрагменти и го експресират на повъ ;рхността си, свързан с молекулите на II HLA клас (DR4, DR1 и в по-слаба степен с DQ2).
II.Медиаторна фаза
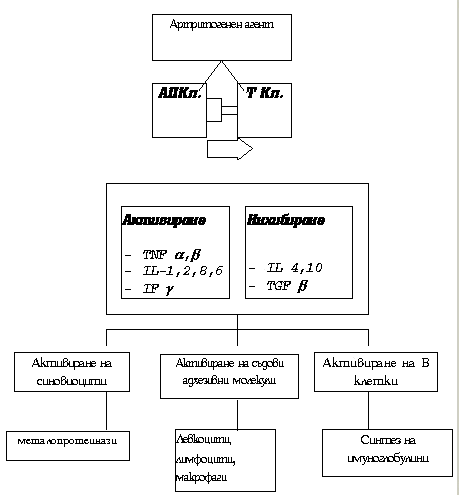
. В ранните етапи на заболяването се наблюдава значително активиране на ендотелните синовиални капилярни клетки, с последваща хиперекспресия на адхезивни мо лекули, увеличаване на левкоцитната миграция от съдовете към екстракапилярните пространства и инфилтрация на синовиалната мембрана с CD4+ Т лимфоцити. Преоблавада производството на цитокини от Th1 т_ 2;п: интерлевкин-2 и g -интерферон (Т хелперите се разделят на две основни субпопулации клетки - Th1 и Th2, като първите при антигенно дразнене секретират интерлевкин-2 и интерферон-g , които ускоряват диференциацията и пролиферацията на Т-цитотоксичните лимфоцити, т.е. усилват клетъчно-медиирания имунитет. Th2 клетките секретират интерлевкин-4, 5, 6 и 10, които стимулират хуморалния иму_ 237;ен отговор чрез ускоряване пролиферацията на В клетките. Съотношението между Th1 и Th2 лимфоц итите има съществена роля в поведението на имунната система при защитата на организма). Последва разраства_ 237;е на синовиалната тъкан с интензивна неоваскуларизация, което довежда до разрушаването на хрущялната тъкан и подлежащата субхондрална кост. Предполага се, че в основата на ревматоидното възпале& #237;ие стои нарушения баланс между провъзпалителните и антиинфламаторните цитокини.Активираните Т лимфоцити синтезират интензивно и в последствие секретират в синовиалната течност и системната циркулация основните проинфламаторни цитокини: интерлевкин 1 и 6 и тумор-некрот изиращ фактор (IL1, IL6, TNF). Именно те инициират първоначалния имунен отговор на организма при острата възпалителна реакция. По-късно към тах се добавят и други медиатори - тромбоцитен растежен фактор, епидер ;мален растежен фактор, фибробластен растежен фактор, инсулино-подобен растежен фактор, невронен растежен фактор и др., които поддържат активността на възпалението, подпомагат неговото хронифицир ;ане и в по-късните етапи участват в костно-хрущялната деструкция, клетъчната пролиферация и фиброза. Това е така, тъй като тези медиатори оказват въздействие преимуществено върху клетките, регулир_ 224;щи костния и хрущялен метаболизъм. Освен това, на системната циркулация на тези фактори се дължат и част от общите прояви при РА.
Цитокините се синтезират няколко часа след първоначалния сигнал за активиране на съответните имунокомпетентни клетки. Приблизително след още толкова се екскретират от синтезиращата ги кл_ 9;тка в екстрацелуларното пространство и се диспергират в съответните биологични течности. Те не са биологично активни в свободно състояние, а изявяват ефекта си само след свързването със съответн 32;те рецептори на прицелната клетка. Силата на въздействието им зависи от продължителността на контакта им с мембранния рецептор.
В отговор на цитокинова синтеза започва синтезата и екскрецията на съответните цитокинови инхибитори. Всъщност това е балансиращата система на организма, оказваща протективен и антиинфлам_ 224;торен ефект.
III. Лимфоидна фаза
. Антиген-представящите клетки активират и хелперните Т лимфоцити. Те стимулират пролиферацията и диференциацията на В лимфоцитите в моноцити за синтез на спе цифични антитела срещу инфекциозния етиологичен фактор. Това е възловият етап в развитието на възпалителната реакция, тъй като от този момент започва трансформацията на процеса в автоимунен.Вирусът на Ebstein-Barr е лимфоцитотропен. Навлизайки в организма той се свързва със съответния рецептор на повърхността на В клетките, прониква в тях и хибридизира своята ДНК молекула с тази на лимфоцит 24;. Новосинтезира се специфичен екстрахируем нуклеарен антиген на РА (RANA). RANA и антителата, насочени към него са открити през 1976 год от Alspaugh и Tan и са доказани с метода на Ochterlony - насрещна дифузия и преципитация в агар. Ин_
4;ектираните клетки започват да произвеждат алотипно-променени имуноглобулини, които в случая играят ролята на автоантигени. Автоантигени се получават и по друг механизъм - специфичните имуноглоб ;улини, насочени към екзогенния етиологичен агент се свързват с него, което обаче променя структурата им и ги превръща в чуждеродни. Имунокомпетентните клетки реагират веднага и срещу RANA и срещу фено 42;ипно-променените имуноглобулинови молекули, изработвайки специфични антитела. Тези от т ях, които са насочени към антигенните детерминанти в константните региони на IgG се наричат ревматоидни факт& #238;ри (РФ). Синтезираните РФ могат да бъдат основно от IgM, IgG и IgA класове и много по-рядко от останалите (IgD, IgE). Освен стимулирането на Т-хелперната актвиност, през тази фаза се наблюдава и подтискане на биологичнат ;а активност на Т-супресорите, което допълнително стимулира синтезата на РФ. Синтезата на РФ е най-типичното имунно нарушение и същевременно една от най-характерните особености на РА. РФ обаче не са с_ 9;ецифични само за РА. Те могат да бъдат открити при почти всички състояния, асоциирани с образуването на имунни комплекси (ИК) или протичащи с хипергамаглобулинемия (хронични бактериални инфекции, за ;болявания с лимфопролиферативен характер, чернодробни заболявания, системни форми на автоимунитет, злокачествени процеси и др., включително и при здрави хора).Въпросът за специфичността на РФ не се изчерпва само със способността им да свързват алотипно различните имуноглобулини (и да формират ИК). До голяма степен РФ имат и специфичност към нуклеарни 42;е антигени и в този смисъл показват качества на анти-нуклеарни антитела. Причината за това се търси във възможните различни механизми, предизвикващи синтезата на РФ - те могат да се синтезират не сам_ 238; под влияние на алотипно променени имуноглобулини, но и от нуклеарни антигени и ИК.
Следва свързването на РФ с прицелните имуноглобулини в ИК. Формирането им е един от най-съществените моменти в патогенезата на артрита. Съществуването на РФ от различни класове на имуноглобули ;ни и възможността образуваните с тях ИК да полимеризират определя формирането на различни по големина, подвижност и разтворимост ИК. В началото на имунната реакция се формират т.нар. междинни ИК, неф ;иксиращи комплемент, които включват комплекса IgG-IgGРФ-комплемент. Тези ИК са по-леки, транспортабилни, склонни към дисеминиране и това е вероятно едно от условията за генерализиране и системни прояви н а патологичния процес. По-нататък тези комплекси се свързват с IgM РФ, образувайки големи, тежки, ИК. Именно те продължават по-нататък патогенетичната верига на ревматоидния възпалителен процес, свърз_ 6;айки големи количества комплемент.
Концентрацията на ИК зависи от степента на образуването (количеството на антигена, активността на синтез на антителата и на комплементарната система) и отстранаяването им (размери, биохимична структура, активност на мононуклеарната фагоцитираща система).
В патогенезата на РА вземат участие и други антитела, а не само РФ. Най-важните са представени в следващата таблица.
|
Вид антитяло |
Прицел |
Патогенетична роля |
Често та (%) |
|
АНА* |
Различни нуклеарни компоненти |
Клетъчна смърт |
4-60 |
|
Антихистони |
Хистони I-IV |
Автоимунни васкулити |
10-30 |
|
Анти РНП** |
РНП |
Възможна последица на поликлонална В кл. Активация. |
30 |
|
Антиперинуклеарни |
Перинуклеарни гранули |
Последица на вирусна инфекция |
70 |
|
Антикератинови |
Кератин |
Тежест на заболяването |
95 |
|
Антикардиолипинови |
Дифосфатидил глицерол |
Ефект върху простагландин-I 2 освобождаване и тромбоцитната агрегация |
10 |
|
Антиколагенови |
Колаген тип II |
Фиксиране на комплемент. Ставни увреди. |
25 |
* анти нуклеарни антитела
** рибонуклеопротеин
IV. Агресивна фаза
. Фагоцитиращите полиморфонуклеари, фибробластите и тъканните фагицити инкорпорират ИК, включват ги в лизозомите си, лизират ги като по този начин ги отстраняв& #224;т от биологичните течности и тъканите. Фагиращите клетки, натоварени с ИК могат да бъдат открити в ставната течност при микроскопско изследване на последната. Това са т.нар. ”рагоцити”. ИК с по-малки ра_ 231;мери се задържат по-дълго време в синовиалната течност и в системното кръвоовбращение в сравнение с тези, имащи средни или големи размери. При фагоцитозата на ИК голяма част от фагиращите клетки за 27;иват и изливат съдържанието на лизозомите си в екстрацелуларното пространство.Активирането на фагоцитиращите клетки в инфламираните стави води до образуване на големи количества високореактивни окисни радикали. При респираторния ”взрив” се генерират супероксидни йо_ 7;и и водороден прекис. Супероксидните йони дифундират слабо от клетките, за разлика от Н
2О2. Последният е изключително мощен оксидант.
Обща схема на патогенезата на РА
АП – антиген-представяща клетка.
V.Деструктивна фаза
. Екстрацелуларните матриксни молекули на съединителната тъкан са подложени на деструктивното действие на много фактори: лизозомни протеолитични ензими, _ 246;итокини, продукти на метаболизма на арахидоновата киселина, свободни кислородни радикали.Свободните кислородни радикали имат многопосочно действие: деполимеризират синовиалната хиалуронидаза, активират латентните металопротеинази, инактивират протеазните инхибитори, гене_ 0;ират липидни пероксиди, които са изключително реактивоспособни, водят до образуването на хемотаксисни простаноиди, увреждат ДНК, водят до апоптоза, инхибират клетъчната метаболитна активнсост, _ 6; частност хрущялната макромолекулна синтеза от хондроцитите.
Стимулираните от цитокините хондроцити освобождават големи количества NO. Чрез него IL1 инхибира протеогликансинтетазата и стимулира металопротеиназната активност. Освен това NO води до увреди 37;а ДНК молекулата, инактивира дихателната верига, води до едем на тъканите, стимулира ангиогенезата и води до хондроцитна апоптоза.
Освободената фосфолипаза в хода на възпалителната реакция разгражда фосфолипидите до арахидонова киселина. Тя от своя страна се подлага на действието на два ензима - циклооксигеназа (с образу ване на простагландини) и липооксигеназа (с последващо образуване на левкотриени). Секрецията на арахидоновите метаболити е до голяма степен регулирано от цитокините. Най-голямо участие във възпа_ 5;ителния процес имат простагландин Е2 и левкотриени А4 и В4. Простагландин Е2 супресира функцията на хелперните лимфоцити, стимулира активацията на плазминогеновия активатор и секрецията на колаген& #224;зата. Левкотриените имат разнообразни проинфламаторни ефекти - индукция на неутрофилен хемотаксис, левкоцитна адхезия към ендотелиите, неутрофилна дегранулация, освобождаване на лизозомни ен_ 1;ими.
В по-ранните етапи се освобождават неутрални протеинази (серинови и металопротеинази). В резултат на действието им се разрушават матриксните макромолекули на хрущяла и прилежащите тъкани, ела стичните влакна на най-малките кръвоносни съдове и хрущялните протеогликани. Към това се добавя и ефекта на трипсина, химиотрипсина и каликреина върху прицелните структури на съединителната тъка н, кръвните белтъци и клетъчните мембрани. Активираната каликреин-кининова система води до вторично задействане на коагулационната каскада, индуцира се агрегация на тромбоцитите, води до освобож даването на серотонин, повишава мембранната пропускливост, стимулира конверсията на С3 фракцията на комплемента в активната й форма С3а. Протеолизата на множество плазмени и тъканни белтъци води д 38; натрупване на множество нискомолекулярни пептиди с изразена биологична активност. Колагеназата лизира колагеновите фибрили, които са практически резистентни на други ензими. В крайна с метка н_ 9;утралните протеази ”потгатвят” съединително
-тъканните матриксни компоненти за действието на киселите хидролази (аспартинови, цистеинови и др.) в по-късните етапи. Разградните съединително-тъканн& #232; продукти се възприемат от имунокомпетентните клетки като чужди антитени, срещу които започва синтез на антитела. Затваря се патологичен цикъл, който поддържа активността на имунната възпалител& #237;а реакция и подпомага нейното хронифициране.Може би най-важните проинфламаторни медиатори са цитокините IL1b и туморният некротизиращ фактор-a (TNFa ). И двата цитокина се произвеждат от макрофагите след стимулиращият ефект на интерферон-g (IFg ) от бактериалните продукти или от комплементфиксиращи ИК. IL1 подпомага лимфоцитното активиране и стимулира макрофагите за производство на протеолитични ензими и хемотаксисни фактори. Същевремен ;но стимулира и остеокластите, което води до костна резорбция. Ревматоидният панус съдържа TNFa в изобилие. Този цитокин също стимулира лимфоцитите и макрофагите, притежава хемоатрактивни свойства и индуцира вторично производство на интерлевкини 6 и 13, които медиират острофазовия отговор. То& #233; също участва в хрущялната и костна деструкция, намалява минералното съдържание на субхондралната кост, като активира остеокластите и благоприятства развитието на остеопорозата и костните еро 31;ии. На мястото на лизираните хрущялни и костни клетки разраства патологична фиброзна тъкан, вследствие на активиране на клетъчната фибробластна пролиферация и ангиогенеза.
Основният прицел на РА са ставите. Всичките по-горе споменати процеси водят до изразено възпаление на ставната синовия, разрушаване на хрущяла, образуването на панус, остеопороза и костни & #229;розии, а в крайните етапи и до анкилоза. Това и определя основната характеристика на РА - ерозивно-деструктивен, прогресиращ полиартрит.
КНИГОПИС
![]()
Home | Home of Mednet | mail to Author
This Site Is Powered By LiveBg.Net Internet Service Provider
(p) 2000 ,Pleven,Bulgaria